|
Clayton Foundation Biochemical lnstitute. Department of Chemistry. The University ofTexas Austin. Texas 78712. USA It appears to me that finally. a glimmcr ofinsight is to be seen in the basic mechanism of oncogcnic transformation. The emcrging picturc is based on fundamental observations from a large number of sources. Wc hayc becn privilcged to scc the results of some ofthc currently brightest rescarch in the mosaic during this workshop. The framcwork for thc cmerging picture is based on thc recognition within recent years that cellular differentiation is a social event among cells in which one cell typc triggers the differentiation of another. This principle hardly would have been a rcvelation to thc early embryologists. Perhaps the foremost. tcchnically accessiblc problem of biology at this time is to undcrstand the specific biochemical mcchanisms by which these processes are implcmented within the cell. The Emerging Field Theory Certain cells excrt their function in inducing differentiation
in target cells eithcr through cell-to-cell physical contact or
from a distance. In both cascs. specific substances. frequently
glycoproteins. from one cell interact with highly specific rccertors
on the target cclls. If the triggcring substance is releascd from
the cell it may be commonly known as a growth substance or hormone.
These factors may be active at chemically incredibly low concentration".
At this mceting Malcolm Moore has rcrorted that lactoferrin acts
as a falctor for thc growth ofcolonies of myeloid cells and that
the activc component is cffective at concentrations of 10 -14M!
For the differentiation falctors to be effective at thesc concentrations.
the binding constant fo)r interaction with the rcccptor must bc
cxtremely largc. In most cases it appears that binding of' no more
than a few moleculcs per target cell is adequate to clicit thc response.
In general. a special systcm will bc required to amplify the signal
rcceived at thc cell surfalcc to a level that it can bc cffective
on intracellular rcactions. It is not clear whether or not part
or all ofthc differentiation factor molecules themsclyes must be
somehow transported to the nuclcus to cffect transcri ptional con
trol. Thc rcla tion of the cell ular com ponen ts in volyed are
schematically derictcd in Fig. I. A proper relation in time diuring
develormcnt as well as space is implicit in the scheme. Differentiation
may occur only during a transient period when cells exhibiting the
appropriate recognition sites arc exposed to the 326 Hardcsty. B
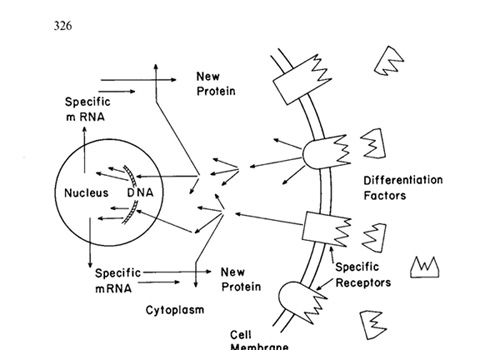
proper signal molecules. Undifferentiated cells may be maintained in the adult organism. then triggered to differentiate into a mature or terminal cell type in which cell division is limited or completely stopped. Differentiation of blood cells provides a classical example of such a system. ThUS the function of the cellular genome during differentiation might be equated more correctly with a library than a computer. Information stored in the DNA is expressed when the proper request is received rather than at a fixed point in a freerunnIng program. Recent evidence appears to indicate that at least in some and probably most situations. the immediate. causal alteration that results in oncogenic transformation involves a lesion in the information retrieval system rather than DNA itself. Theoretically the lesion might occur at any point from the differentiation factors themselves to the synthesis of a new protein. A number ofspecific mechanisms can be envisioned. I divide them into four classes based on the location of the lesion : I. Defective or Deficient Differentiation Factors A differentiating cell might not be exposed to the proper factors, perhaps because oftime-space relationships with other cells or because ofa lesion in the cells producing the appropriate differentiation factor(s). In this case the oncogenic cell might be normal in its capacity to recognize and respond to differentiation factors which are not present in its environment. Certain teratocarcinoma appear to be excellent candidates for this class. Exciting experiments involving implantation of normal embryonic and teratocarcinoma cells into pseudo-pregnant mice to form normal, chimeric or allophenic progeny strongly support such a model (K. Illmensee, and B. Mintz, 1976). Hopefully, additional successful experiments in this area will be reported shortly. II. Defective Recognition Sites Another theoretically possible situation might involve numerically inadequate or defective recognition sites on the target cell. Chemically inert, surface reactive materials such as asbestos may function as carcinogens by this mechanism. Such cells might be induced to differentiate if the concentration of the critical factors could be elevated to a level at which they could induce a nonproliferative state. There are a number of reports involving leukemias that may reflect this situation (L. Sachs, 1974; R. C. Gallo et al., 1977) or possible Type I defects. Certain neuroblastomas that can be stimulated to differentiate with cAMP (K. N. Prasad and A.W. Hsie, 1971) may fall into this category. Neuroblastomas that can be stimulated to differentiate in the presence ofglia cells (C. P. Reynolds and J. R. Perez-Polo, 1975) and pheochromocytomas in which neurite proliferation can be stimulated with nerve growth factor (A. S. Tischler and L. A. Greene, 1975) might involve lesions at the level of either the differentiation factor or the cellular receptor. III. Defective Intracellular Signal Transmission The next level of lesion that can be envisioned involves transmission of the signal received at the cell surface to the transcriptional and translational machinery of the cell. Some virus-induced transformation appears to provide an example of a lesion at this level. Rat kidney cells that have been transformed by Rous sarcoma virus can be caused to revert to a normal phenotype by inhibiting protein synthesis, presumably by blocking the synthesis of an unstable product of the transforming gene (J.F. Ash et al., 1976). The src gene appears to code for a protein kinase that may interfere with intracellular control mechanisms as mentioned below and considered by Ray Erikson at this meeting. It appears that avian and murine leukemia virus carry transforming genes that are quite different from the src gene and code for other proteins that may disrupt the intracellular transmission of the signal in different ways. Murine erythroid cells transformed by Friend leukemia virus provide a system that may belong to this class. Differentiation can bc induced in these cells by the simple expedient ofgrowing them in tissue culture in the presence of dimethylsulfooxide (C. Friend et al., 1971) or a number ofother aprotic solven ts. In all of the examples given above the transformed cells can be induced to undergo differentiation if they arc exposedi to the proper external stimulus. The block in differentiation can be overcome! In effect, the cells can be cured of their "transformed" condition! The point of fundamental importance is that transformation has not involved an irreversible loss of genetic information or a structural change in DNA that precludes further differentiation. IV. Defective Malignant Cell DNA Yet another class of oncogenic transformation may involve structural alterations of DNA so that the af fected cells lack genetic information required foor terminal differentiation. Certain genetically determined cancers may fall into this class. However. it should be noted that some imaginal disc tumors of Drosophila that clearly are inherited according to the principles ofMendelian genetics may revert to a differentiating state (E. Gateff , 1978a). Genetically inherited imaginal disc tumors fall into two classes. THose that appear to be irreversibly transformed and others that are capable ofdifferentiation when they develop in close contact with wild type cells (E. Gateff, 1978 b). It appears likely that the latter class may involve a mutation that affects the production of a differentiation substance produced in non-malignant cells and thus probably should be classified as a Type I transformation. X-ray induced leukemias may provide another example ofa Type IV transformation in which radiation has resulted in damage of genes required fo)r a late stage of differentiation which has occurred in an undifferentiated cell type. The damage may not be expressed for a relatively long time until the defective cells are induced to start along a differentiation pathway. Thus transfo)rmation resulting from changes in the DNA of the malignant cells fat 11 into a fundamentally different category than the three classes considered above. in that they involve seemingly irreversible loss of essential genetic information. Session Highlights Peter Duesberg Ray Erikson Gisela Kramer Ian Kerr Gebhard Koch The Future I believe any detailed prediction of future developments in molecular biologv and biochemistry are likely to be wrong or at least incomplete, probably to a major extent. However, there are key problems that must receive continual consideration. and several areas that appear to be ripe for investigation. The concepts inherent in the mechanism of normal differentiation reflected in Fig. I will be tested and retested in many systems in the forthcoming years. The most crucial problem for an understanding of normal differentia tion is the molecular mechanism(s) by which signals received at the cell surface activate transcription from specific genes. Is part or all of the protein differentiation factor taken into the cell and used in the activation process itself perhaps at the DNA level as appears to be the case with steroid hormones. or are intermediate reactions involved? For most systems, the physical and chemical characterization of the differentiation factors and their specific receptors presents a severe technical problem in working with the very small quantities that are available. In many cases the assay systems used to monitor isolation are not quantitative and are no more than marginally satisfactory. This presents a formidable problem, especially when the biological response depends on two or more specific components, as frequently appears to be the case. The development of better assay systems. especially in vitro systems involving specific biochemical reactions rather than the response of intact cells. is critical to satisfactory progress in this area. Wi th respect to the seq uence of in tracell ular reactions triggered by growth substances. two problems or areas stand out as being both technically feasible and crucially important. The first problem involves the mechanism by which signals received at the cell surface are amplified and transmitted to target reactions in the cytoplasm and nucleus. It appears that there are likely to be a number of alternative mechanisms to cascade systems involving protein kinases for amplification of the signal received at the cell surface. The small nucleotide described by Ian Kerr that is produced as part of the interferon and double-stranded RNA system seems to be part of such a system. It is likely that other types of amplification mechanisms will be found. The second problem involves regulation of cAMP-independent protein kinases. A number and perhaps a great many cAMP-independent protein kinases may be involved in amplification and transmission of cell surface signals. The enzyme system that is activated by double-stranded RNA and interferon is an excellent example. What is the specific molecular mechanism by which such enzymes are activated and do they function in cascade sequences? Wi th respect to transformed cells, the search for differen tia tion factors and conditions with which transformed cells can be induced to either stop dividing or differentiate to a non-dividing form appears to be the key problem. However it is frought with technical limitations that may limit progress until they are resolved. The transforming genes and their products are ripe for investigation and an investigative effort will be made in a number of laboratories. Are transforming genes really cellular genes that have been integrated into a viral genome in such away that they no longer respond to the normal control systems within the cell? What is the biochemical mechanism by which the product of transforming genes disrupt differentiation and induce the physiological changes associated with transformation? It appears possible that the next Wilsede workshop may include hard answers to some of these problems and a consideration of substantiated models of the molecular mechanism by which oncogenic transformation occurs. Eventually, I believe such insight will provide the basis tor a rational therapy to cure leukemia at the cellular level. References Ash. J F. Vogt P K Singcr S .I Rcyersion from transformed to mrmal
phenotype by inhibition of protein synthesis in rat kidncv cells
infected with a tempcraturc-scnsitivc mutant of Rous sarcoma virus
Proc Nat Acad Sci USA 73, 3603-3607 ( 1976 ) |